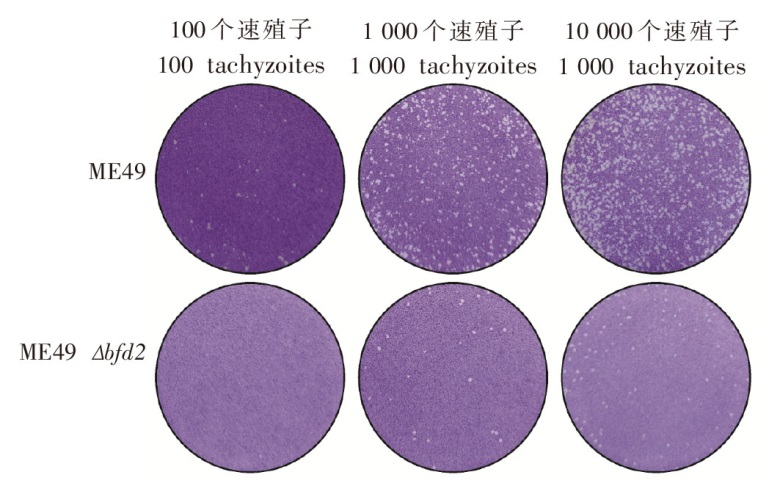
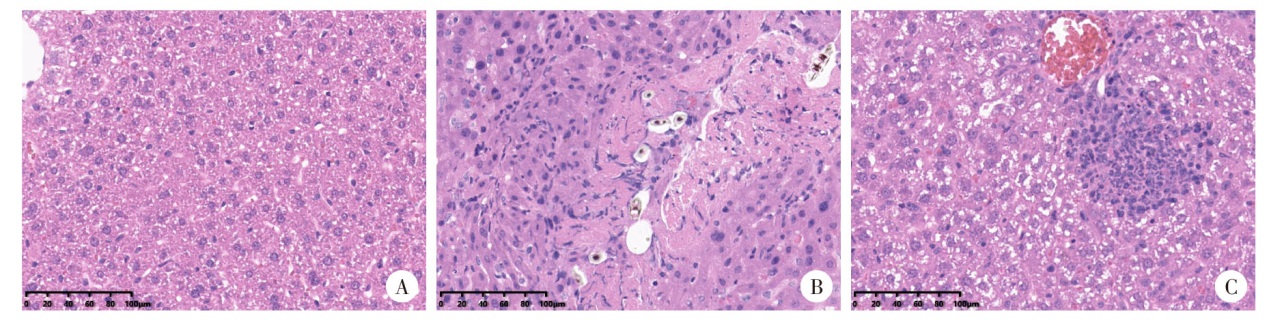
| [1] | Atif I, Touloun O, Boussaa S. Toxoplasma gondii in humans, animals and in the environment in Morocco: A literature review[J]. Gut Pathog, 2024, 16(1):53. | | [2] | Hussain MA, Stitt V, Szabo EA, et al. Toxoplasma gondii in the food supply[J]. Pathogens, 2017, 6(2): 21. | | [3] | Sgroi G, Viscardi M, Santoro M, et al. Genotyping of Toxoplasma gondii in wild boar (Sus scrofa) in southern Italy: Epidemiological survey and associated risk for consumers[J]. Zoonoses Public Health, 2020, 67(7): 805-813. | | [4] | Deng HF, Swart A,Bona?i? Marinovi? AA, et al. The effect of salting on Toxoplasma gondii viability evaluated and implemented in a quantitative risk assessment of meat-borne human infection[J]. Int J Food Microbiol, 2020, 314: 108380. | | [5] | Murillo-Léon M, Bastidas-Quintero AM, Steinfeldt T. Decoding Toxoplasma gondii virulence: The mechanisms of IRG protein inactivation[J]. Trends Parasitol, 2024, 40(9): 805-819. | | [6] | Zhao XY, Ewald SE. The molecular biology and immune control of chronic Toxoplasma gondii infection[J]. J Clin Invest, 2020, 130(7): 3370-3380. | | [7] | Pittman KJ, Aliota MT, Knoll LJ. Dual transcriptional profiling of mice and Toxoplasma gondii during acute and chronic infection[J]. BMC Genomics, 2014, 15(1): 806. | | [8] | Diniz DG, Guerreiro LCF, et al. Contrasting disease progression, microglia reactivity, tolerance, and resistance to Toxoplasma gondii infection in two mouse strains[J]. Biomedicines, 2024, 12(7): 1420. | | [9] | Tyumentseva M, Tyumentsev A, Akimkin V. CRISPR/Cas9 landscape: Current state and future perspectives[J]. Int J Mol Sci, 2023, 24(22): 16077. | | [10] | 王聪, 程维晟, 刘芳, 等. 基于CRISPR/Cas9技术的弓形虫rop16Ⅰ/Ⅲ缺陷虫株的构建及毒力鉴定[J]. 中国人兽共患病学报, 2017, 33(1): 22-26, 31. | | | Wang C, Cheng WS, Liu F, et al. CRISPR/Cas9-based construction of rop16Ⅰ/Ⅲ deficient strain of Toxoplasma gondii and its virulence identification[J]. Chin J Zoonoses, 2017, 33(1): 22-26, 31. (in Chinese) | | [11] | 吴燕, 张欣, 李瑾, 等. 基于CRISPR/Cas9技术的弓形虫病疫苗研究进展[J]. 中国血吸虫病防治杂志, 2024, 36(5): 542-547. | | | Wu Y, Zhang X, Li J, et al. Progress of researches on toxoplasmosis vaccines based on the CRISPR/Cas9 technology[J]. Chin J Schisto Control, 2024, 36(5): 542-547. (in Chinese) | | [12] | 贾永根, 闫爱霞, 黄敏君, 等. 基于CRISPR/Cas9技术对刚地弓形虫假定蛋白TGGT1_310420的研究[J]. 中国寄生虫学与寄生虫病杂志, 2019, 37(2): 150-154, 160. | | | Jia YG, Yan AX, Huang MJ, et al. CRISPR/Cas9-based localization and functional analysis of Toxoplasma gondii putative protein TGGT1_310420[J]. Chin J Parasitol Parasit Dis, 2019, 37(2): 150-154, 160. (in Chinese) | | [13] | Seo HH, Han HW, Lee SE, et al. Modelling Toxoplasma gondii infection in human cerebral organoids[J]. Emerg Microbes Infect, 2020, 9(1): 1943-1954. | | [14] | Wu MM, Cudjoe O, Shen JL, et al. The host autophagy during Toxoplasma infection[J]. Front Microbiol, 2020, 11: 589604. | | [15] | Licon MH, Giuliano CJ, Chan AW, et al. A positive feedback loop controls Toxoplasma chronic differentiation[J]. Nat Microbiol, 2023, 8(5): 889-904. | | [16] | Wang FR, Holmes MJ, Hong HJ, et al. Translation initiation factor eIF1.2 promotes Toxoplasma stage conversion by regulating levels of key differentiation factors[J]. Nat Commun, 2024, 15(1): 4385. | | [17] | Waldman BS, Schwarz D, WadsworthMH 2nd, et al. Identification of a master regulator of differentiation in Toxoplasma[J]. Cell, 2020, 180(2): 359-372. e16. | | [18] | Walrad P, Paterou A, Acosta-Serrano A, et al. Differential trypanosome surface coat regulation by a CCCH protein that co-associates with procyclin mRNA cis-elements[J]. PLoS Pathog, 2009, 5(2): e1000317. | | [19] | Wang JL, Li TT, Elsheikha HM, et al. The protein phosphatase 2A holoenzyme is a key regulator of starch metabolism and bradyzoite differentiation in Toxoplasma gondii[J]. Nat Commun, 2022, 13(1): 7560. | | [20] | Gupta D, Bhattacharjee O, Mandal D, et al. CRISPR-Cas9 system: A new-fangled dawn in gene editing[J]. Life Sci, 2019, 232: 116636. |
|

 )(
)( ), HU Yuan1, PENG Hui1, SHEN Yujuan1, LIU Hua1, CAO Jianping1,2,*(
), HU Yuan1, PENG Hui1, SHEN Yujuan1, LIU Hua1, CAO Jianping1,2,*(