中国寄生虫学与寄生虫病杂志 ›› 2026, Vol. 44 ›› Issue (2): 222-228.doi: 10.12140/j.issn.1000-7423.2026.02.011
朱弟( )(
)( ), 吴蔚玲, 孔德豪, 周志豪, 彭鸿娟*()()
), 吴蔚玲, 孔德豪, 周志豪, 彭鸿娟*()()
ZHU Di()(), WU Weiling, KONG Dehao, ZHOU Zhihao, PENG Hongjuan*()()
摘要:
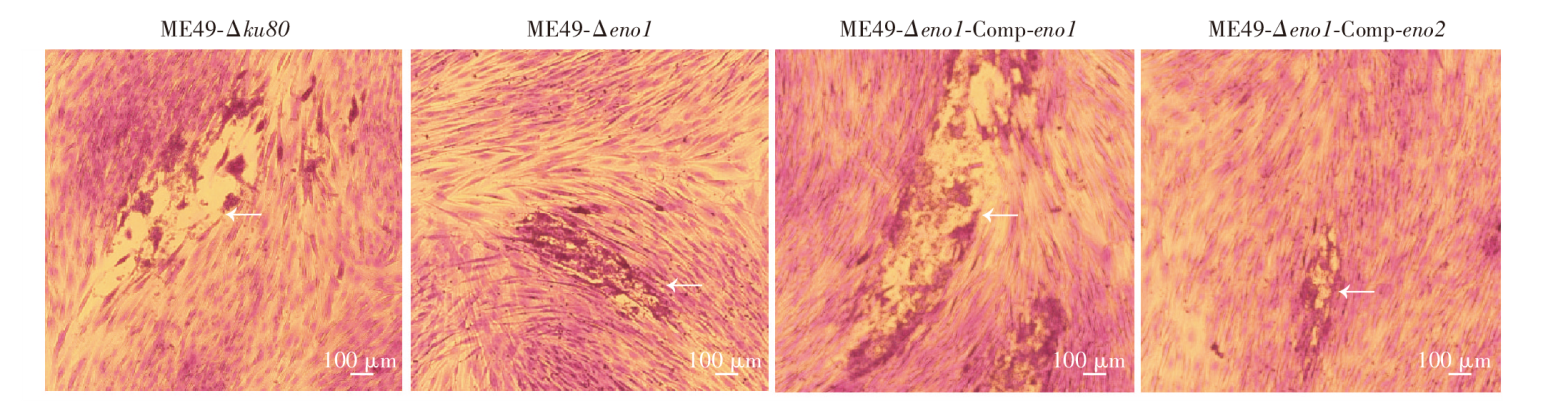
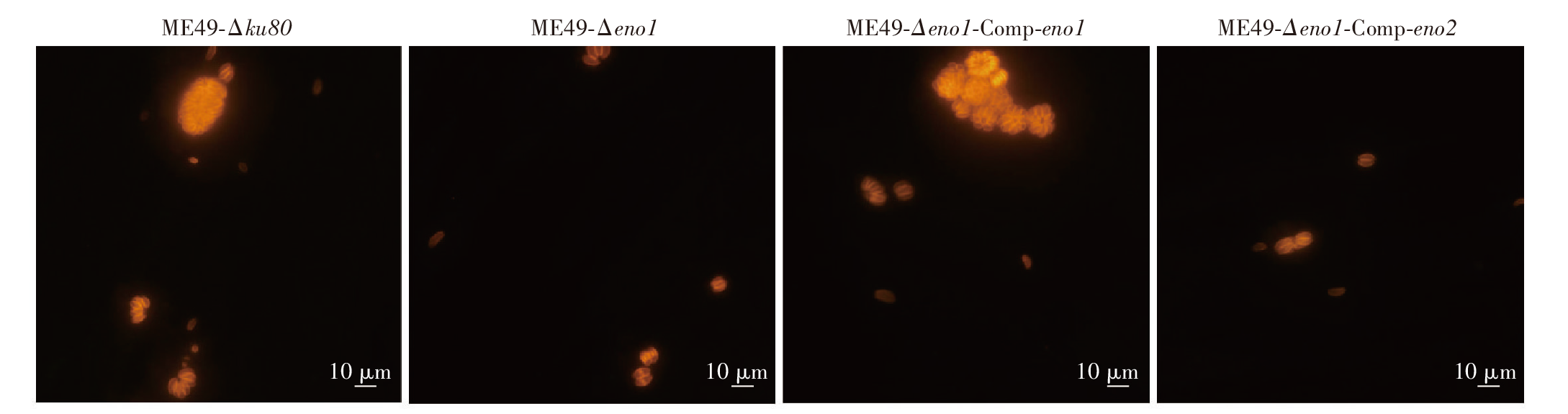
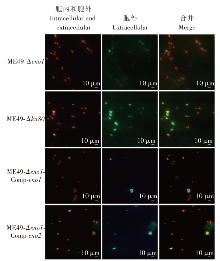
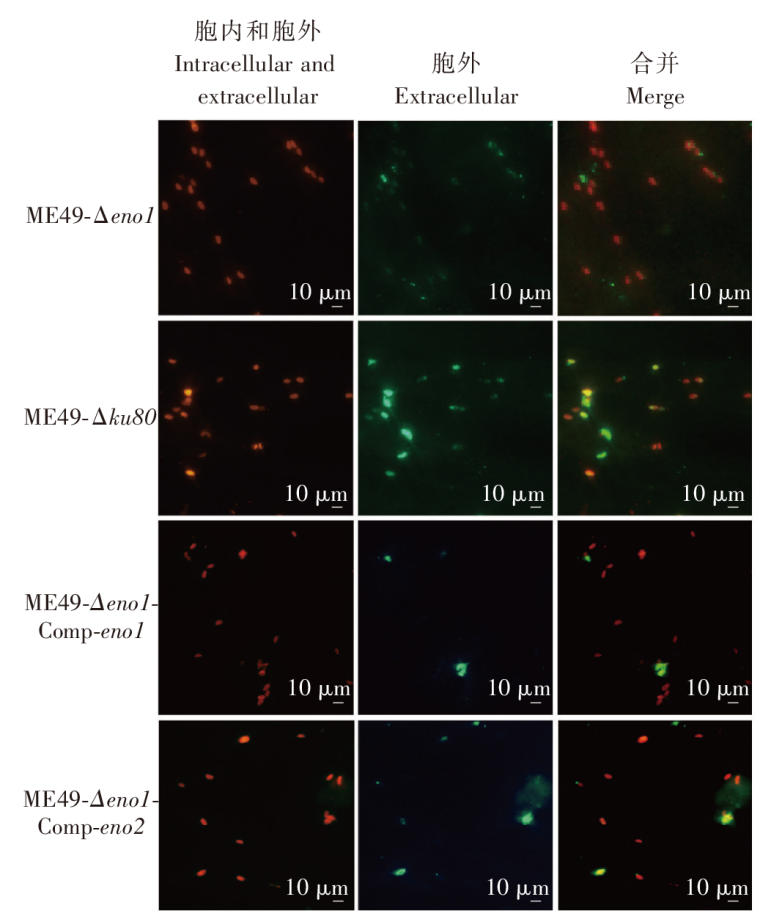
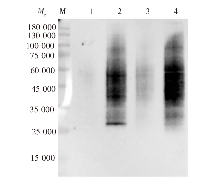
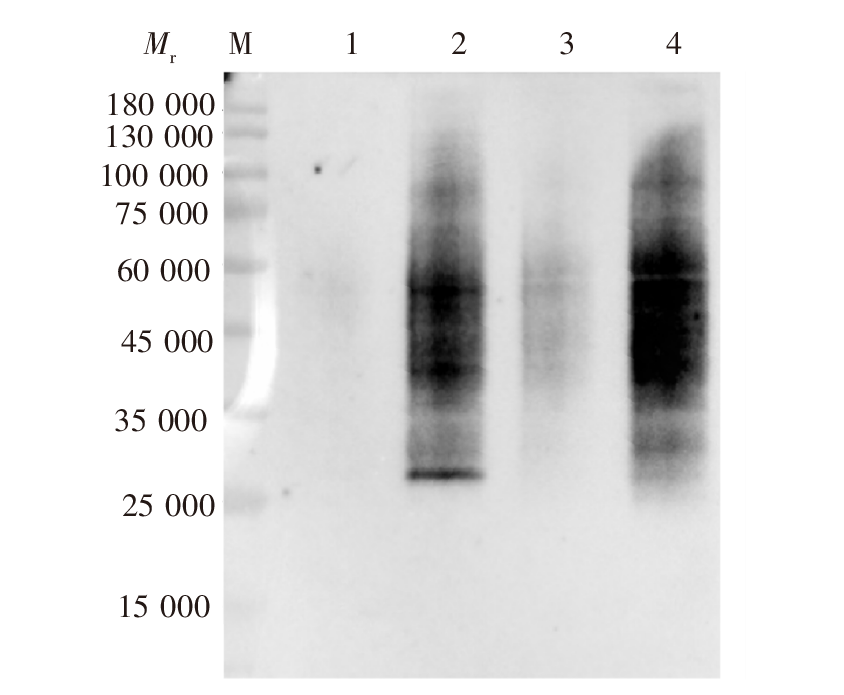
目的 探讨刚地弓形虫缓殖子阶段特异性烯醇酶1(ENO1)在速殖子与缓殖子相互转化过程中的生物学功能,分析ENO家族在弓形虫期转化调控网络中的作用及分子机制。方法 利用成簇规律间隔短回文重复序列及相关蛋白9(CRISPR/Cas9)基因编辑技术构建eno1敲除虫株ME49-Δeno1及ME49-Δeno1-Comp-eno1和ME49-Δeno1-Comp-eno2两种回补虫株;采用空斑实验、增殖实验和入侵实验,比较ME49-Δku80、ME49-Δeno1、ME49-Δeno1-Comp-eno1和ME49-Δeno1-Comp-eno2等4种虫株感染人包皮成纤维细胞后的生长、增殖和入侵能力。采用体外碱性诱导后,实时荧光定量PCR(qPCR)检测ME49-Δku80、ME49-Δeno1速殖子期特异性基因表面抗原1(sag1)和缓殖子期特异性基因缓殖子抗原1(bag1)的相对转录水平,评价eno1敲除对弓形虫速殖子向缓殖子转化能力的影响。采用嘌呤霉素标记与蛋白质免疫印迹(Western blotting)检测ME49-Δku80和ME49-Δeno1两种虫株新生肽链合成情况,验证eno1敲除对弓形虫蛋白质合成的影响。结果 成功构建了ME49-Δeno1虫株,以及ME49-Δeno1-Comp-eno1与ME49-Δeno1-Comp-eno2两种回补虫株。空斑实验、增殖实验和入侵实验结果显示,ME49-Δku80、ME49-Δeno1、ME49-Δeno1-Comp-eno1和ME49-Δeno1-Comp-eno2虫株形成的空斑面积分别为(16 538 ± 14 310)、(4 376 ± 5 355)、(17 377 ± 14 333)和(8 710 ± 8 207)μm2,平均每个纳虫泡的弓形虫速殖子数量分别为(5.836 ± 0.382)、(4.792 ± 0.150)、(5.165 ± 1.243)和(3.872 ± 0.301)个,入侵效率分别为(57.802 ± 6.932)%、(19.679 ± 4.508)%、(63.833 ± 1.198)%和(35.901 ± 8.098)%,eno1敲除抑制了弓形虫的生长(F = 38.290,P < 0.01)、胞内增殖(F = 4.467,P < 0.05)及入侵能力(F = 36.650, P < 0.01)。eno1的回补完全恢复了弓形虫的生长等上述能力,ME49-Δeno1-Comp-eno1虫株形成的空斑面积、纳虫泡内平均速殖子数量和入侵效率均与ME49-Δku80虫株相比差异无统计学意义(t = 0.459、0.895、1.485,P > 0.05)。eno2回补部分恢复了弓形虫的生长等能力,ME49-Δeno1-Comp-eno2虫株形成的空斑面积大于ME49-Δeno1虫株(t = 5.263,P < 0.05),但小于ME49-Δku80虫株(t = 4.905,P < 0.01);纳虫泡内平均速殖子数量少于ME49-Δku80(t = 6.998,P < 0.05);入侵效率高于ME49-Δeno1虫株(t = 3.032,P < 0.05),但低于ME49-Δku80虫株(t = 3.559,P < 0.05)。碱性条件诱导48 h后,ME49-Δeno1虫株sag1基因的相对转录水平为1.067 ± 0.115,与ME49-Δku80株的1.002 ± 0.154相比差异无统计学意义(t = 0.584,P > 0.05);ME49-Δeno1虫株bag1基因的相对转录水平为13.172 ± 1.679,低于ME49-Δku80株的20.338 ± 1.344(t = 5.770,P < 0.01)。碱性诱导96 h后,ME49-Δeno1虫株sag1基因的相对转录水平为0.957 ± 0.132,高于ME49-Δku80的0.727 ± 0.053(t = 2.803,P < 0.05);ME49-Δeno1虫株bag1基因的相对转录水平为162.248 ± 23.377,低于ME49-Δku80的231.413 ± 12.910(t = 4.486,P < 0.05)。Western blotting检测结果显示,ME49-Δku80和ME49-Δeno1虫株在新生肽链合成水平上无明显差异,eno1敲除对弓形虫蛋白质合成能力无明显影响。结论 缓殖子特异性烯醇酶ENO1促进弓形虫速殖子生长,并正向调控速殖子向缓殖子的转化,是该过程中的关键调控分子,但并非必需分子,而且ENO1在弓形虫速殖子增殖中不可被ENO2所替代。
中图分类号: