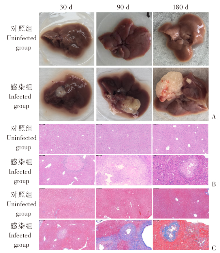
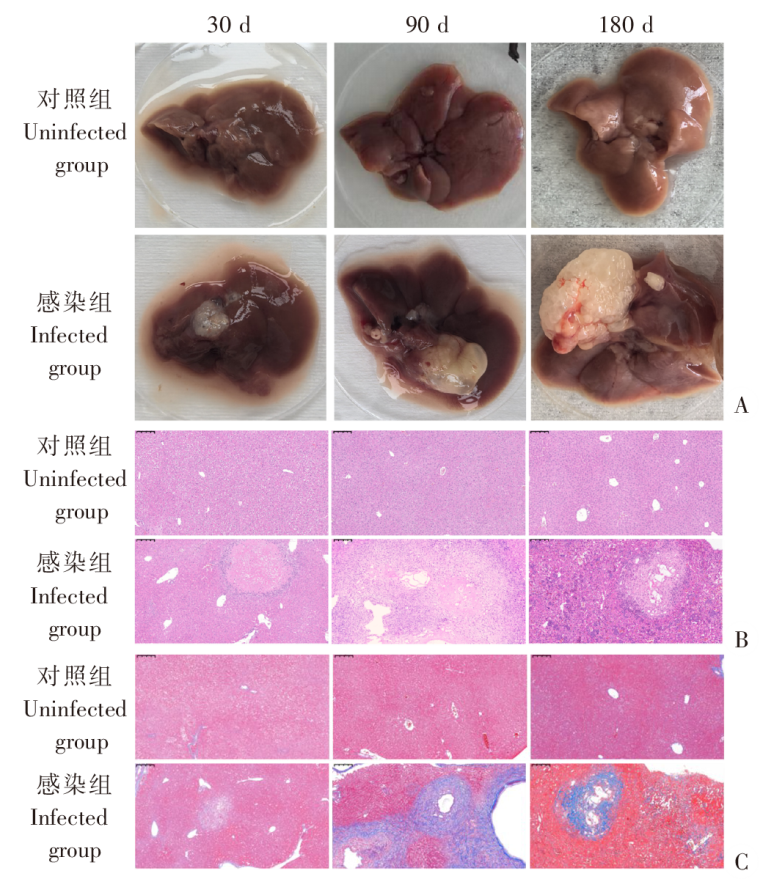
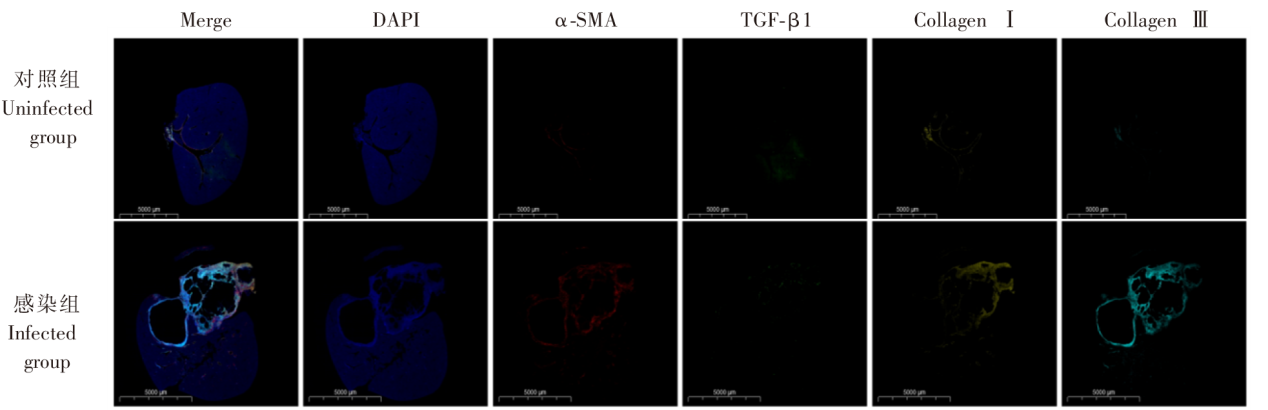
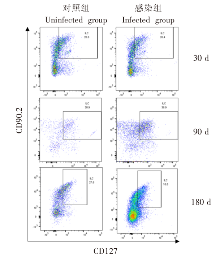
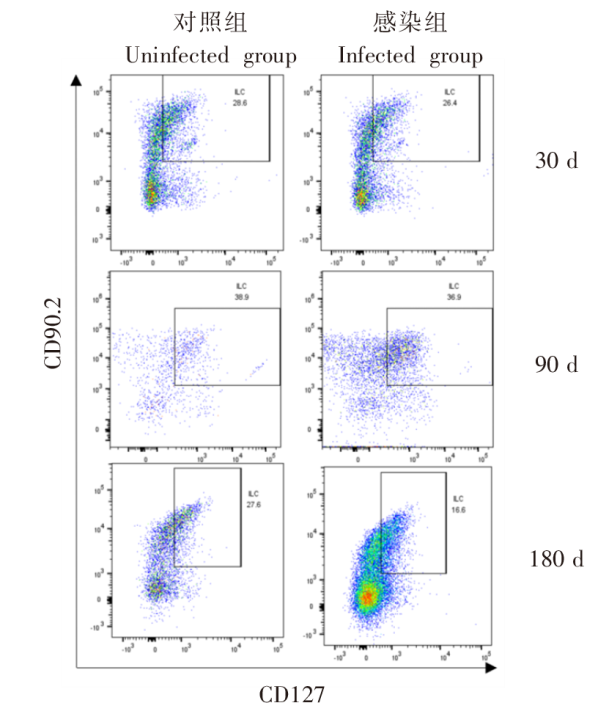
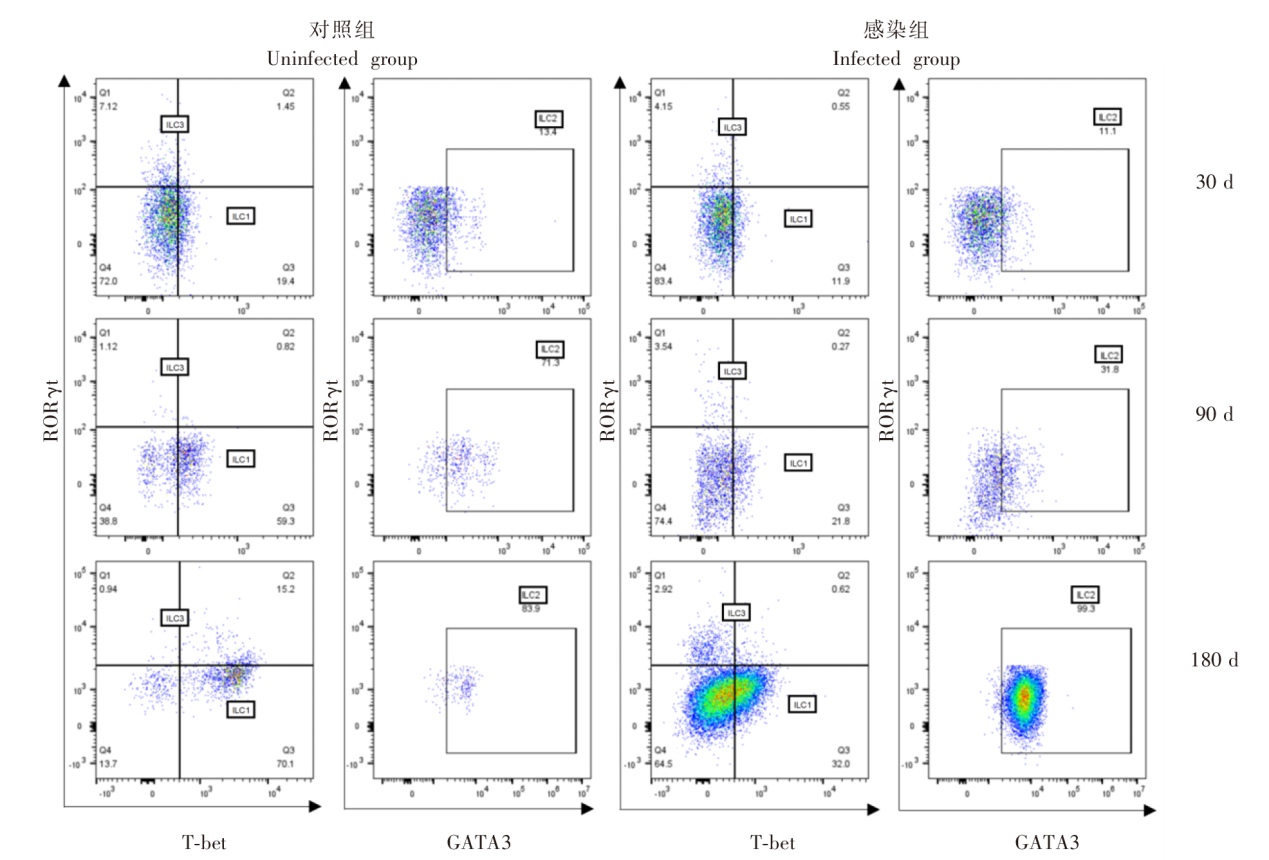
| [1] | Lundstr?m-Stadelmann B, Rostami A, Frey CF, et al. Human alveolar echinococcosis-global, regional, and national annual incidence and prevalence rates[J]. Clin Microbiol Infect, 2025, 31(7): 1139-1145. | | [2] | Li J, Yang YY, Han XM, et al. Oral delivery of anti-parasitic agent-loaded PLGA nanoparticles: enhanced liver targeting and improved therapeutic effect on hepatic alveolar echinococcosis[J]. Int J Nanomedicine, 2023, 18: 3069-3085. | | [3] | Liu TL, Pu GT, Wang LQ, et al. LncRNA gm40262 promotes liver fibrosis and parasite growth through the gm40262-miR-193b-5p-TLR4/Col1α1 axis[J]. mBio, 2025, 16(4): e0228724. | | [4] | Wang LY, Qin M, Gavotte L, et al. Societal drivers of human echinococcosis in China[J]. Parasit Vectors, 2022, 15(1): 385. | | [5] | 刘白雪, 蒉嫣, 薛垂召, 等. 2023年全国棘球蚴病防治工作进展[J]. 中国寄生虫学与寄生虫病杂志, 2025, 43(1): 8-16. | | | Liu BX, Kui Y, Xue CZ, et al. Progress of the national echinococcosis control programme in China, 2023[J]. Chin J Parasitol Parasit Dis, 2025, 43(1): 8-16. (in Chinese) | | [6] | Wang H, Zhang CS, Fang BB, et al. Dual role of hepatic macrophages in the establishment of the Echinococcus multilocularis metacestode in mice[J]. Front Immunol, 2020, 11: 600635. | | [7] | 王苗苗, 安秀青, 周鸿乾, 等. 肝多房棘球蚴病导致肝硬化的研究进展[J]. 中国人兽共患病学报, 2023, 39(11): 1130-1135. | | | Wang MM, An XQ, Zhou HQ, et al. Research progress in hepatic alveolar echinococcosis leading to cirrhosis[J]. Chin J Zoonoses, 2023, 39(11): 1130-1135. (in Chinese) | | [8] | 徐凯, 王海久, 张丽, 等. 多房棘球蚴对宿主肝细胞损害机制的研究进展[J]. 中国寄生虫学与寄生虫病杂志, 2021, 39(2): 256-260. | | | Xu K, Wang HJ, Zhang L, et al. Research progress on the mechanisms underlying the impairment of host hepatocytes by Echinococcus multilocularis[J]. Chin J Parasitol Parasit Dis, 2021, 39(2): 256-260. (in Chinese) | | [9] | 司晶, 唐露, 李臣诚, 等. 肝纤维化的发病机制及其治疗药物研究进展[J]. 药学进展, 2021, 45(3): 205-211. | | | Si J, Tang L, Li CC, et al. Pathogenesis of liver fibrosis and progress of research on its related therapeutic drugs[J]. Prog Pharm Sci, 2021, 45(3): 205-211. (in Chinese) | | [10] | Dai MY, Yang XY, Yu YH, et al. Helminth and host crosstalk: new insight into treatment of obesity and its associated metabolic syndromes[J]. Front Immunol, 2022, 13: 827486. | | [11] | Nono JK, Lutz MB, Brehm K. Expansion of host regulatory T cells by secreted products of the tapeworm Echinococcus multilocularis[J]. Front Immunol, 2020, 11: 798. | | [12] | 张涛, 张耀刚, 杨紫晗, 等. 多房棘球蚴原头节感染后阻断PD-1对巨噬细胞吞噬功能的影响[J]. 中国人兽共患病学报, 2023, 39(11): 1095-1099. | | | Zhang T, Zhang YG, Yang ZH, et al. Effects of PD-1 blockade on macrophage phagocytosis after Echinococcus multilocularis protoscoleces infection[J]. Chin J Zoonoses, 2023, 39(11): 1095-1099. (in Chinese) | | [13] | Kabata H, Motomura Y, Kiniwa T, et al. ILCs and allergy[J]. Adv Exp Med Biol, 2022, 1365: 75-95. | | [14] | Leupold T, Wirtz S. ILCs-crucial players in enteric infectious diseases[J]. Int J Mol Sci, 2022, 23(22): 14200. | | [15] | Srivastava RK, Sapra L, Bhardwaj A, et al. Unravelling the immunobiology of innate lymphoid cells (ILCs): implications in health and disease[J]. Cytokine Growth Factor Rev, 2023, 74: 56-75. | | [16] | 赵童, 徐成灵, 李高斌, 等. ILC2在2型炎症型哮喘中的作用[J]. 中国医学创新, 2024, 21(9): 163-168. | | | Zhao T, Xu CL, Li GB, et al. The role of ILC2 in type 2 inflammation in asthma[J]. Med Innov China, 2024, 21(9): 163-168. (in Chinese) | | [17] | 于明凯, 朱玥洁, 丁剑冰, 等. Ⅱ型固有淋巴细胞在肝脏疾病中的作用研究进展[J]. 中华肝脏病杂志, 2023, 31(7): 776-780. | | | Yu MK, Zhu YJ, Ding JB, et al. Research progress on the role of type Ⅱ innate lymphoid cells in liver diseases[J]. Chin J Hepatol, 2023, 31(7): 776-780. (in Chinese) | | [18] | Toki S, Goleniewska K, Zhang J, et al. TSLP and IL-33 reciprocally promote each other’s lung protein expression and ILC2 receptor expression to enhance innate type-2 airway inflammation[J]. Allergy, 2020, 75(7): 1606-1617. | | [19] | Hildreth AD, Padilla ET, Tafti RY, et al. Sterile liver injury induces a protective tissue-resident cDC1-ILC1 circuit through cDC1-intrinsic cGAS-STING-dependent IL-12 production[J]. Cell Rep, 2023, 42(2): 112141. | | [20] | Curio S, Belz GT. The unique role of innate lymphoid cells in cancer and the hepatic microenvironment[J]. Cell Mol Immunol, 2022, 19(9): 1012-1029. | | [21] | Liu Y, Liu MR, Zhang CM, et al. The silencing of NREP aggravates OA cartilage damage through the TGF-β1/Smad2/3 pathway in chondrocytes[J]. J Orthop Translat, 2024, 44: 26-34. | | [22] | 张伶慧, 陈根, 种世桂, 等. 多房棘球蚴病中免疫细胞调控机制的研究进展[J]. 中国寄生虫学与寄生虫病杂志, 2022, 40(1): 109-113, 120. | | | Zhang LH, Chen G, Chong SG, et al. Research progress on the immune regulation mechanism in alveolar echinococcosis[J]. Chin J Parasitol Parasit Dis, 2022, 40(1): 109-113, 120. (in Chinese) | | [23] | 张靖仪, 田凤鸣, 姜涛, 等. 单核细胞源性巨噬细胞MAC387对肝泡型包虫病患者肝脏炎症与纤维化的作用[J]. 新发传染病电子杂志, 2023, 8(1): 19-25. | | | Zhang JY, Tian FM, Jiang T, et al. The role of macrophage MAC387 in the inflammation and fibrosis of hepatic alveolar echinococcosis of human cases[J]. Electron J Emerg Infect Dis, 2023, 8(1): 19-25. (in Chinese) | | [24] | Wen TH, Tsai KW, Wu YJ, et al. The framework for human host immune responses to four types of parasitic infections and relevant key JAK/STAT signaling[J]. Int J Mol Sci, 2021, 22(24): 13310. | | [25] | Esperante D, Gutiérrez MIM, Issa ME, et al. Similarities and divergences in the metabolism of immune cells in cancer and helminthic infections[J]. Front Oncol, 2023, 13: 1251355. | | [26] | Steinmann S, Schoedsack M, Heinrich F, et al. Hepatic ILC2 activity is regulated by liver inflammation-induced cytokines and effector CD4+ T cells[J]. Sci Rep, 2020, 10(1): 1071. | | [27] | 张亚光, 孙兵. 二型免疫细胞(Th2/ILC2)在肺部过敏性炎症中作用机制的研究进展[J]. 中国免疫学杂志, 2024, 40(1): 11-20. | | | Zhang YG, Sun B. Progress on mechanism of type Ⅱ immune cells (Th2/ILC2) in allergic pulmonary inflammation[J]. Chin J Immunol, 2024, 40(1): 11-20. (in Chinese) | | [28] | Wu XF, Ming BX, Wu T, et al. IL-33/ST2 axis contributes to the dermal fibrosis of systemic sclerosis via promoting fibroblasts activation[J]. J Dermatol Sci, 2022, 107(2): 95-104. | | [29] | Cosway EJ, James KD, White AJ, et al. The alarmin IL33 orchestrates type 2 immune-mediated control of thymus regeneration[J]. Nat Commun, 2023, 14(1): 7201. | | [30] | Weiskirchen R, Tacke F. Interleukin-33 in the pathogenesis of liver fibrosis: alarming ILC2 and hepatic stellate cells[J]. Cell Mol Immunol, 2017, 14(2): 143-145. | | [31] | Cheng SL, Ma XM, Wu BJ, et al. Therapeutic effects of IL-33/ST-2 pathway inhibition combined with albendazole on hepatic fibrosis and immune regulation in alveolar echinococcosis: in vivo and in vitro evidence[J]. Parasit Vectors, 2026, 19(1): 177. | | [32] | 赵涵玥, 李锦田, 薛俊隆, 等. 多房棘球蚴感染晚期小鼠肝脏中NK1.1的表达对T细胞功能的影响[J]. 中国寄生虫学与寄生虫病杂志, 2024, 42(5): 594-600, 607. | | | Zhao HY, Li JT, Xue JL, et al. Effect of NK1.1 expression on liver T cell function in the advanced stage of mice infected with Echinococcus multilocularis[J]. Chin J Parasitol Parasit Dis, 2024, 42(5): 594-600, 607. (in Chinese) | | [33] | 孜比姑·肉素, 阿比旦·艾尼瓦尔, 阿迪莱·多力坤, 等. LAG3缺陷对多房棘球蚴感染小鼠自然杀伤细胞功能及肝纤维化的影响[J]. 中国血吸虫病防治杂志, 2024, 36(1): 59-66. | | | Zibigu RS, Abidan ANWE, Adilai DLK, et al. Effect of LAG3 deficiency on natural killer cell function and hepatic fibrosis in mice infected with Echinococcus multilocularis[J]. Chin J Schisto Control, 2024, 36(1): 59-66. (in Chinese) | | [34] | Savage TM, Fortson KT, de Los Santos-Alexis K, et al. Amphiregulin from regulatory T cells promotes liver fibrosis and insulin resistance in non-alcoholic steatohepatitis[J]. Immunity, 2024, 57(2): 303-318.e6. |
|

 ), SU Yaxin, WANG Ying, ZHOU Hao, CUI Lijun, JIANG Nan, ZHANG Jing, WANG Xiangqing, CAO Jianping, JIANG Bin, HUO Lele*(
), SU Yaxin, WANG Ying, ZHOU Hao, CUI Lijun, JIANG Nan, ZHANG Jing, WANG Xiangqing, CAO Jianping, JIANG Bin, HUO Lele*( )(
)(