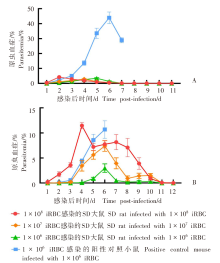
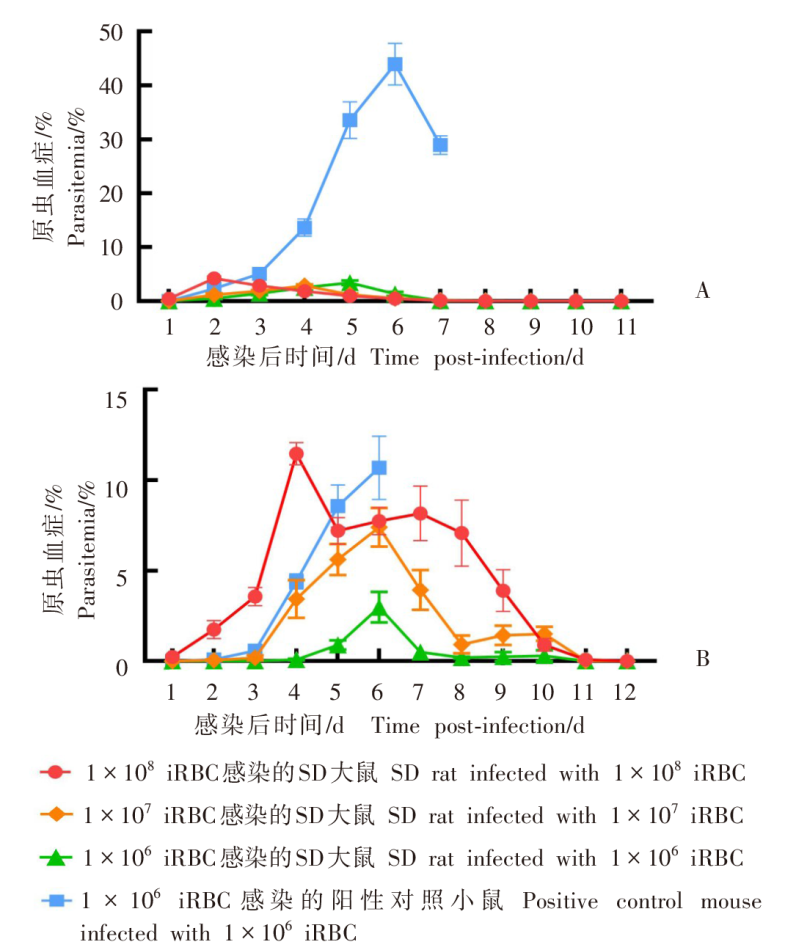
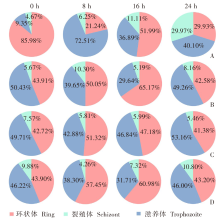
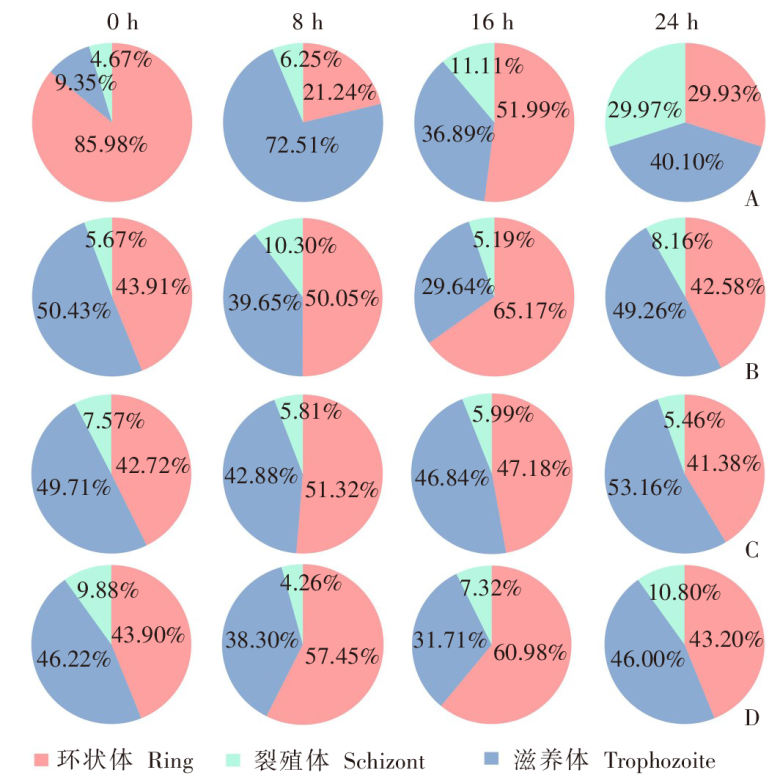
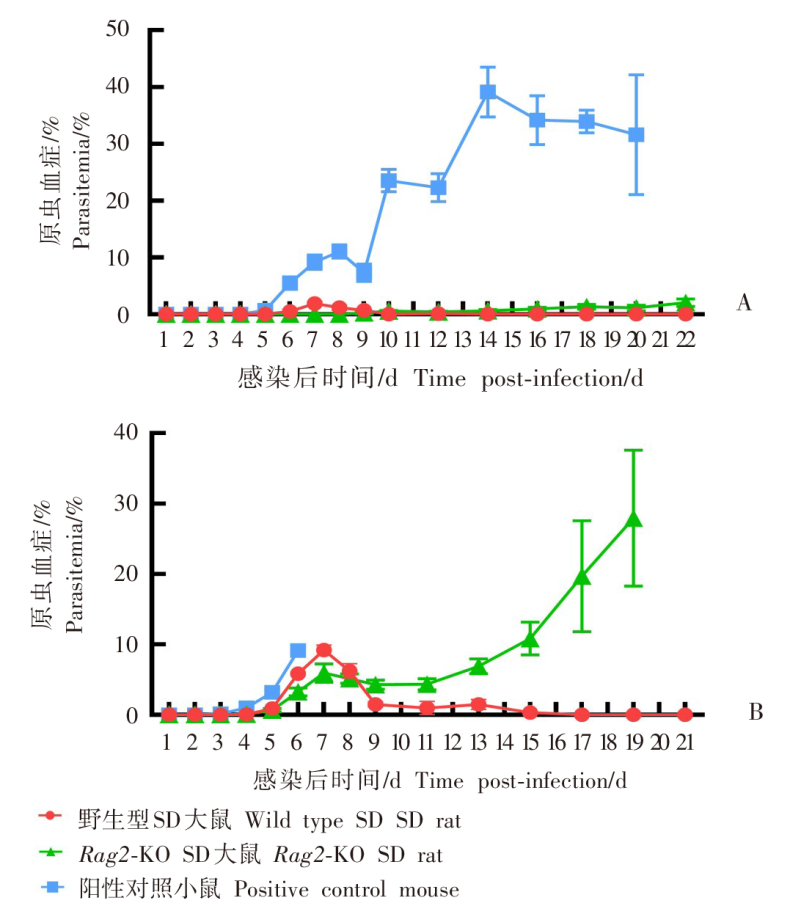
| [1] | World Health Organization. World malaria report 2022[R]. Geneva: WHO, 2022: 14-17. | | [2] | Duffy PE, Patrick Gorres J. Malaria vaccines since 2000: progress, priorities, products[J]. NPJ Vaccines, 2020, 5(1): 48. | | [3] | Klotz FW, Chulay JD, Daniel W, et al. Invasion of mouse erythrocytes by the human malaria parasite, Plasmodium falciparum[J]. J Exp Med, 1987, 165(6): 1713-1718. | | [4] | Tyagi RK, Tandel N, Deshpande R, et al. Humanized mice are instrumental to the study of Plasmodium falciparum infection[J]. Front Immunol, 2018, 9: 2550. | | [5] | Shears MJ, Seilie AM, Kim Lee Sim B, et al. Quantification of Plasmodium knowlesi versus Plasmodium falciparum in the rhesus liver: implications for malaria vaccine studies in rhesus models[J]. Malar J, 2020, 19(1): 313. | | [6] | Zhang JX, Lin BY, Pan YR. Study on the exoerythrocytic forms of Plasmodium yoelii yoelii in rats and mice[J]. Chin J Parasitol Parasit Dis, 1990, 8(4): 288-290. (in Chinese) | | | (张家埙, 林宝英, 潘玉蓉. 约氏疟原虫感染大鼠和小鼠的红外期研究[J]. 寄生虫学与寄生虫病杂志, 1990, 8(4): 288-290.) | | [7] | (AcrouteDVA, Lafitte S, Dive D, et al. A role for CD4+ and CD8+ cells and not for CD25+ cells in the control of Plasmodium berghei ANKA blood stage parasites in rats[J]. Parasite, 2010, 17(1): 53-60. | | [8] | Safeukui I, Gomez ND, Adelani AA, et al. Malaria induces anemia through CD8+ T cell-dependent parasite clearance and erythrocyte removal in the spleen[J]. mBio, 2015, 6(1): e02493-e02414. | | [9] | Cheviet T, Lefebvre-Tournier I, Wein S, et al. Plasmodium purine metabolism and its inhibition by nucleoside and nucleotide analogues[J]. J Med Chem, 2019, 62(18): 8365-8391. | | [10] | Pohl K, Cockburn I A. Innate immunity to malaria: the good, the bad and the unknown[J]. Front Immunol, 2022, 13: 914598. | | [11] | Belachew EB. Immune response and evasion mechanisms of Plasmodium falciparum parasites[J]. J Immunol Res, 2018, 2018: 6529681. | | [12] | Tham WH, Healer J, Cowman AF. Erythrocyte and reticulocyte binding-like proteins of Plasmodium falciparum[J]. Trends Parasitol, 2012, 28(1): 23-30. | | [13] | Kim I, Yang DJ, Tang XY, et al. Reference gene validation for qPCR in rat carotid body during postnatal development[J]. BMC Res Notes, 2011, 4: 440. | | [14] | Schussek S, Groves PL, Apte SH, et al. Highly sensitive quantitative real-time PCR for the detection of Plasmodium liver-stage parasite burden following low-dose sporozoite challenge[J]. PLoS One, 2013, 8(10): e77811. | | [15] | Liehl P, Zuzarte-Luís V, Chan J, et al. Host-cell sensors for Plasmodium activate innate immunity against liver-stage infection[J]. Nat Med, 2014, 20(1): 47-53. | | [16] | Ploemen IHJ, Prudêncio M, Douradinha BG, et al. Visualisation and quantitative analysis of the rodent malaria liver stage by real time imaging[J]. PLoS One, 2009, 4(11): e7881. | | [17] | Borges da Silva H, Fonseca R, Cassado AD, et al. In vivo approaches reveal a key role for DCs in CD4+ T cell activation and parasite clearance during the acute phase of experimental blood-stage malaria[J]. PLoS Pathog, 2015, 11(2): e1004598. | | [18] | Galen SC, Borner J, Martinsen ES, et al. The polyphyly of Plasmodium: comprehensive phylogenetic analyses of the malaria parasites (order Haemosporida) reveal widespread taxonomic conflict[J]. R Soc Open Sci, 2018, 5(5): 171780. | | [19] | Wang XX, Huang FS. Effects of cyclophosphamide on the development of exo-erythrocytic forms of Plasmodum yoelii in rats[J]. Chin J Parasitol Parasit Dis, 1987, 5(3): 186-189. (in Chinese) | | | (王兴相, 黄复生. 环磷酰胺对大鼠体内约氏疟原虫红外期发育的影响[J]. 中国寄生虫学与寄生虫病杂志, 1987, 5(3): 186-189.) | | [20] | Cêtre C, Pierrot C, Cocude C, et al. Profiles of Th1 and Th2 cytokines after primary and secondary infection by Schistosoma mansoni in the semipermissive rat host[J]. Infect Immun, 1999, 67(6): 2713-2719. | | [21] | Boyle MJ, Wilson DW, Richards JS, et al. Isolation of viable Plasmodium falciparum merozoites to define erythrocyte invasion events and advance vaccine and drug development[J]. Proc Natl Acad Sci USA, 2010, 107(32): 14378-14383. | | [22] | Shinkai Y, Rathbun G, Lam KP, et al. RAG-2-deficient mice lack mature lymphocytes owing to inability to initiate V(D)J rearrangement[J]. Cell, 1992, 68(5): 855-867. | | [23] | Noto FK, Adjan-Steffey V, Tong M, et al. Sprague dawley Rag2-null rats created from engineered spermatogonialstem cells are immunodeficient and permissive to human xenografts[J]. Mol Cancer Ther, 2018, 17(11): 2481-2489. | | [24] | Mercado TI. Paralysis associated with Plasmodium berghei malaria in the rat[J]. J Infect Dis, 1965, 115(5): 465-472. | | [25] | Adam E, Pierrot C, Lafitte S, et al. The age-related resistance of rats to Plasmodium berghei infection is associated with differential cellular and humoral immune responses[J]. Int J Parasitol, 2003, 33(10): 1067-1078. | | [26] | Keita Alassane S, Nicolau-Travers ML, Menard S, et al. Young Sprague Dawley rats infected by Plasmodium berghei: a relevant experimental model to study cerebral malaria[J]. PLoS One, 2017, 12(7): e0181300. |
|

 ), HE Biao, GAO Yuanli, FAN Yongling, ZHU Feng, DING Yan, LIU Taiping, XU Wenyue*(
), HE Biao, GAO Yuanli, FAN Yongling, ZHU Feng, DING Yan, LIU Taiping, XU Wenyue*(