中国寄生虫学与寄生虫病杂志 ›› 2023, Vol. 41 ›› Issue (5): 539-545.doi: 10.12140/j.issn.1000-7423.2023.05.003
郭帅( ), 何彪, 高源利, 范永铃, 朱锋, 丁艳, 刘太平, 徐文岳*()
), 何彪, 高源利, 范永铃, 朱锋, 丁艳, 刘太平, 徐文岳*()
GUO Shuai(), HE Biao, GAO Yuanli, FAN Yongling, ZHU Feng, DING Yan, LIU Taiping, XU Wenyue*()
摘要:
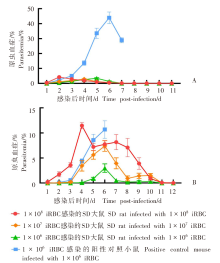
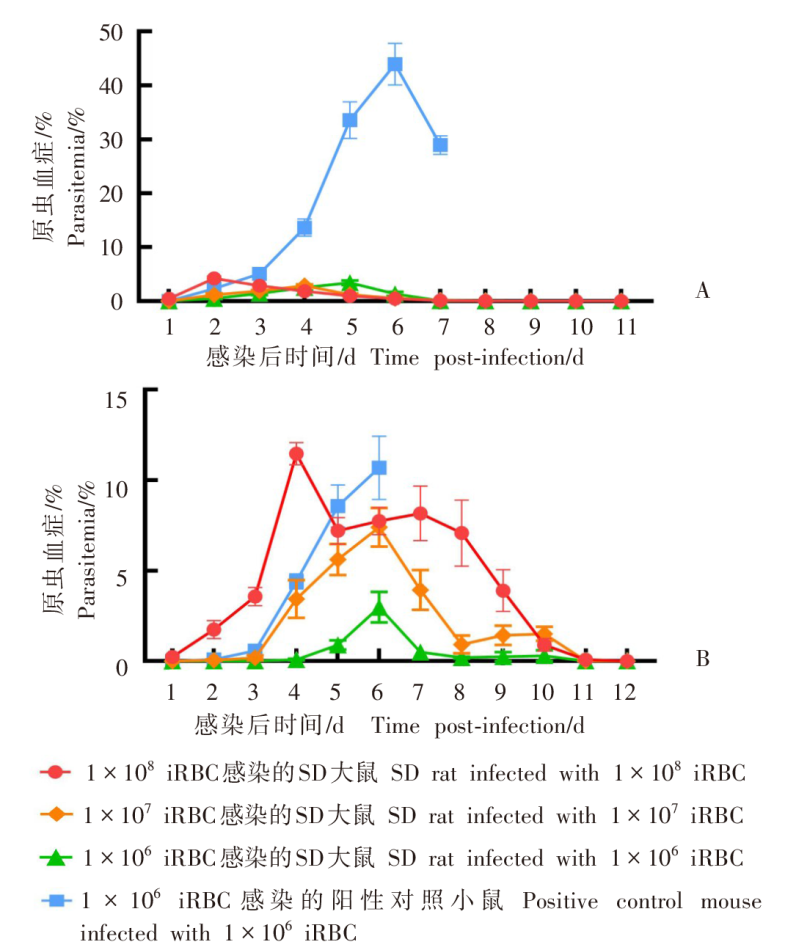
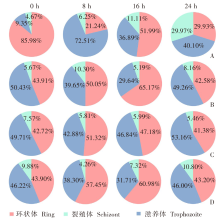
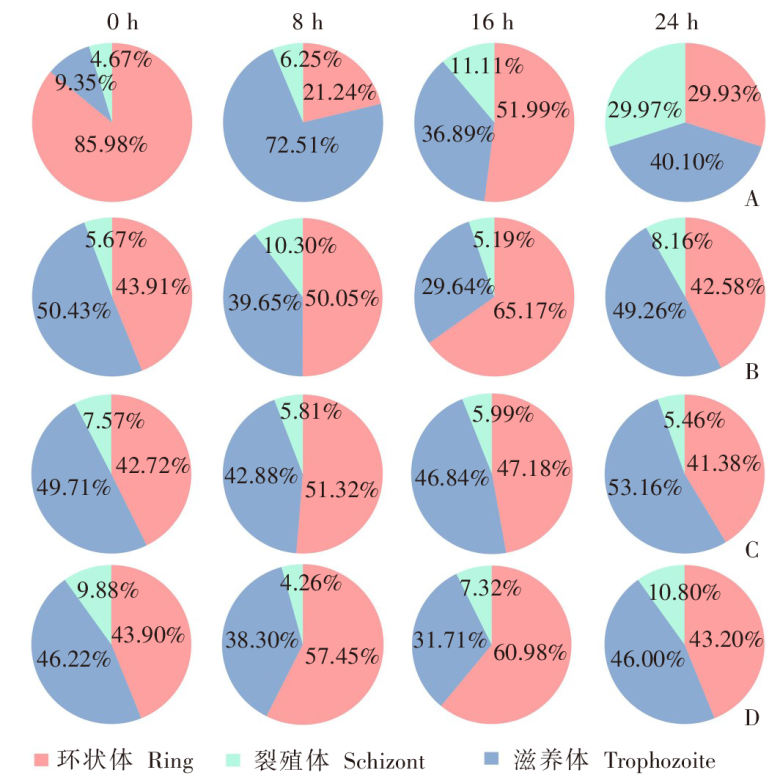
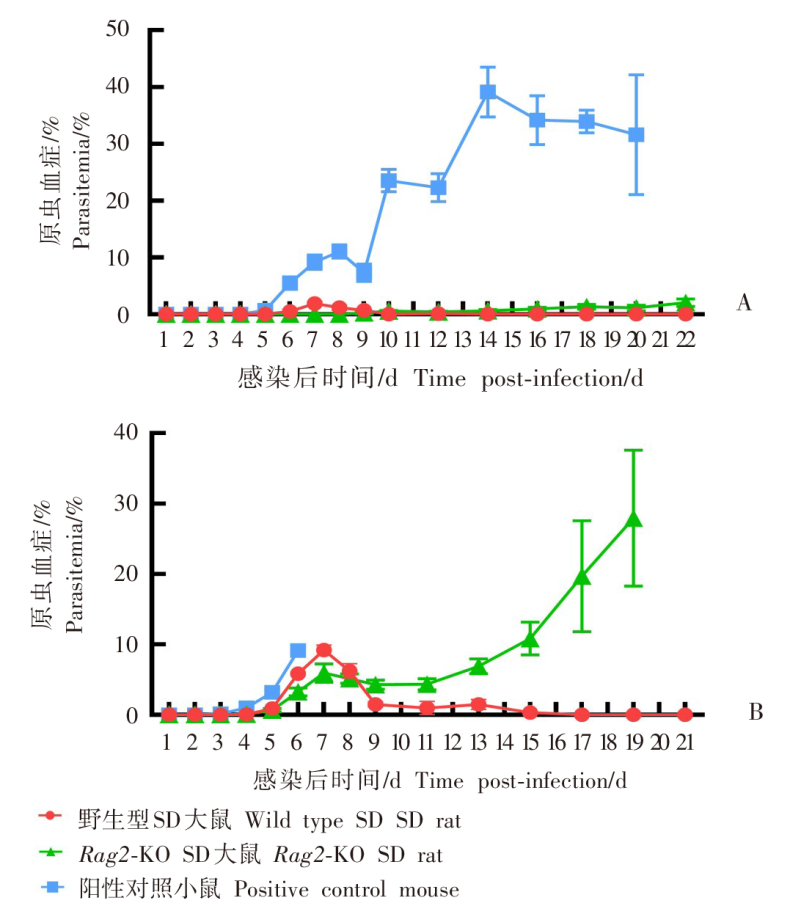
目的 用约氏疟原虫(P.y)BY265株和伯氏疟原虫(P.b)ANKA-luciferase株感染小鼠和大鼠,分析鼠疟原虫感染宿主的种特异性。 方法 制备P.y和P.b的子孢子和裂殖子。分别用5 × 104的P.y和P.b子孢子经尾静脉注射感染SD大鼠、BALB/c小鼠(P.y感染组,5只/组)和SD大鼠、C57BL/6J小鼠(P.b感染组,5只/组),每日取尾静脉血涂片镜检,记录红内期疟原虫出现的时间。P.y子孢子感染SD大鼠和BALB/c小鼠后42 h取肝组织,实时荧光定量PCR(qRT-PCR)检测肝组织疟原虫18S rRNA的相对表达量。P.b子孢子感染SD大鼠和C57BL/6J小鼠后42 h活体成像法检测肝脏荧光值。分别用1 × 108、1 × 107、1 × 106的P.y和P.b被感染红细胞(iRBC)感染SD大鼠(P.y感染组、P.b感染组)和易感小鼠(BALB/c、C57BL/6J)(阳性对照组),每日取尾静脉血涂片镜检,计算原虫血症。分别用P.y、P.b裂殖子感染SD大鼠和易感小鼠,每日取尾静脉血涂片镜检,计算原虫血症;当大鼠、小鼠原虫血症达到峰值后,每隔8小时取尾静脉血涂片镜检,持续24 h,分析疟原虫发育节律;观察大鼠、小鼠实验性脑型疟(ECM)的发生情况。分别用P.y、P.b裂殖子感染T、B细胞缺陷的Rag2-KO SD大鼠、野生型SD大鼠和易感小鼠,每日取尾静脉血涂片观察,计算原虫血症。两组数据之间比较采用非配对t检验,组间原虫血症趋势比较采用双因素方差分析(two-way ANOVA)。 结果 P.y感染组的SD大鼠、BALB/c小鼠,P.b感染组的SD大鼠、C57BL/6J小鼠体内疟原虫均能完成肝期发育,于第3天进入红内期。qRT-PCR结果显示,P.y子孢子感染SD大鼠、BALB/c小鼠体内疟原虫特异性18S rRNA相对表达量分别为(1.63 ± 0.381)、(1.00 ± 0.232),二者差异有统计学意义(t = 2.801,P < 0.05)。活体成像结果显示,P.b子孢子感染SD大鼠体内荧光度值为(6.243 ± 1.425)× 107,高于C57BL/6J小鼠的(1.624 ± 0.530)× 107(t = 6.077,P < 0.01)。红内期iRBC感染结果显示,3种剂量的P.y感染组SD大鼠的原虫血症趋势[峰值分别为(3.500 ± 1.042)%、(2.850 ± 0.627)%、(3.400 ± 0.962)%]之间差异无统计学意义(F = 0.145,P > 0.05),但与阳性对照组[峰值为(43.928 ± 9.448%)]差异有统计学意义(F = 84.040、63.760、58.400,均P < 0.01);P.b感染组SD大鼠的原虫血症趋势[峰值分别为(11.468 ± 1.362)%、(7.398 ± 2.387)%、(2.984 ± 1.881)%]随感染剂量降低而下降,其中1 × 108、1 × 106剂量组原虫血症趋势与阳性对照组[峰值为(10.682 ± 4.278)%]差异有统计学意义(F = 13.83、17.320,均P < 0.01),1 × 107剂量组原虫血症趋势与阳性对照组差异无统计学意义(F = 2.234,P > 0.05)。裂殖子感染结果显示,感染后6 d,P.y感染组SD大鼠、BALB/c小鼠原虫血症分别为(0.902 ± 0.235)%、(17.420 ± 4.105)%,二者差异有统计学意义(t = 9.943,P < 0.01);P.b感染组SD大鼠、C57BL/6J小鼠原虫血症分别为(6.804 ± 2.978)%、(9.290 ± 1.055)%,二者差异无统计学意义(t = 1.759,P > 0.05);P.b感染组SD大鼠、C57BL/6J小鼠ECM累计发生率分别为11/15、13/15,二者差异无统计学意义(t = 1.414,P > 0.05)。发育节律分析结果显示,P.y感染组SD大鼠发育节律与BALB/c小鼠不同,未呈现24 h规律;P.b感染组SD大鼠发育节律与C57BL/6J小鼠相近,具有24 h规律。感染后18 d,P.y感染组Rag2-KO SD大鼠、野生型SD大鼠、BALB/c小鼠原虫血症分别为(1.326 ± 0.908)%、0、(33.937 ± 3.453)%,Rag2-KO SD大鼠与野生型SD大鼠之间原虫血症差异有统计学意义(t = 2.267,P < 0.05);感染后17 d,P.b感染组Rag2-KO SD大鼠、野生型SD大鼠的原虫血症为(19.685 ± 5.752)%、(0.007 ± 0.013)%(t = 2.499,P < 0.05),阳性对照组仅存的1只C57BL/6J小鼠的原虫血症为25.410%。 结论 P.y和P.b子孢子均能感染大鼠并完成肝期发育进入红内期。鼠疟原虫不易感染大鼠的影响因素在红内期,大鼠体内鼠疟原虫可被清除;P.y对宿主种属表现出更强的选择性;P.b感染大鼠的急性期原虫血症和ECM发生率与小鼠差异均无统计学意义。适应性免疫在大鼠彻底清除体内鼠疟原虫中发挥重要作用。
中图分类号: